
Mycobacterial Lsr2 and E. coli H-NS: Structural Basis for Binding AT-Rich Sequences
The mycobacterial Lsr2 story is a curious one. Lsr2 showed up on the scene as an antigen from Mycobacterium leprae back in an Infection and Immunity paper published in 19911. Some work was done to sequence the antigen’s gene and the efforts of the 90’s revolved around leprosy and the immune response. Not much else was done for a while. If you poke around with BLAST, you will discover that lsr2 is well conserved in mycobacteria and actinomycetes, thus it is potentially interesting outside the realm of leprosy. Around 2006 to 2008, it was found that if you delete lsr2 in M. smegmatis it disrupts the ability to form biofilms2, changes the colony morphology from rough to smooth2 and results in a hypermotile phenotype3. Around the same time, publications came out that Lsr2 was involved in transcriptional regulation of multi-drug tolerance and antibiotic-induced responses4, as well as protecting against reactive oxygen intermediates5, in M. tuberculosis! This was intriguing, but the underlying molecular mechanisms still remained unknown.
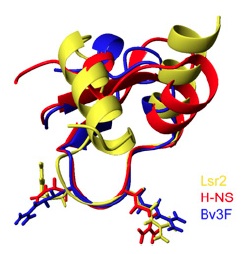
It is at this point, around 2008, that Jun Liu’s Lab (University of Toronto) started to shed some light on the mechanistic basis of Lsr2. They were able to show that M. tuberculosis Lsr2 is able to bridge distant segments of DNA6, much like the histone-like nuleoid structuring protein H-NS in E. coli. They could even complement an E. coli H-NS defect using M. tuberculosis Lsr27, suggesting that Lsr2 is an analogue of H-NS. This was exciting stuff, since Lsr2 and H-NS have no sequence homology and based on structural comparisons, the connection would not have likely been made. In their most recent articles (Gordon et al., 2010 and 2011), they used ChIP-on-chip analysis to perform high-resolution, genome-wide mapping of Lsr2 binding sites revealing that Lsr2 binds many genes involved in M. tuberculosis virulence and immunogenicity, including the ESX secretion system and PE/PPE genes. They also determined the tertiary structure of the Lsr2 DNA-binding domain, showing that the Arg97-Gly98-Arg99 residues are inserted into the minor groove of DNA at AT-rich regions. This is similar to the AT-hook motif in mammalian non-histone chromatin protein HMGA, which also binds AT-rich DNA and is involved in chromatin structure and gene regulation. Other residues in Lsr2 interact with the sugar-phosphate backbone on either edge of the minor groove and increase binding affinity. They then went on to determine the structural basis for recognition of AT-rich DNA by other unrelated silencing proteins, namely H-NS and Bv3F. In their 2011 paper,they show that it is the width of the minor grove that determines the binding preference of H-NS and Lsr2. The figure above (Fig. 3F from Gordon et al. 2011) shows the superimposed structures of Salmonella H-NS (red), Burkholderia Bv3F (blue) and Mycobacterium Lsr2 (yellow). You can see that the loop consisting of the conserved “T/SXQ/RGRXPA” motif of these proteins adopts nearly identical conformations. This is an excellent example of convergent evolution and definitely some Good Reads!
- Sela, S. (1991) Identification of Mycobacterium leprae antigens from a cosmid library: characterization of a 15-Kilodalton antigen that is recognized by both the humoral and cellular immune systems in leprosy patients. Infection and Immunity 59: 4117-4124.
- Chen, J.M., et al. (2006) Roles of Lsr2 in colony morphology and biofilm formation of Mycobacterium smegmatis. J. of Bacteriology 188: 633-641.
- Arora, K., et al. (2008) Inactivation of lsr2 results in a hypermotile phenotype in Mycobacterium smegmatis. J. of Bacteriology 190: 4291-4300.
- Colangeli, R., et al. (2007) Transcriptional regulation of multi-drug tolerance and antibiotic-induced responses by the histone-like protein Lsr2 in M. tuberculosis. PLoS Pathogens 3: 780-793.
- Colangeli, R., et al. (2009) The multifunctional histone-like protein Lsr2 protects mycobacteria against reactive oxygen intermediates. PNAS 106:4414-4418.
- Chen, J.M., et al. (2008) Lsr2 of Mycobacterium tuberculosis is a DNA-bridging protein. Nucleic Acids Research 26:2123-2135.
- Gordon, B.R., et al. (2008) Lsr2 of Mycobacterium represents a novel class of H-NS-like proteins. J. of Bacteriology 190:7052-7059.
