
The Evolving Story of Phage λ Transcriptional Control: Enhancer-like Transcriptional Activation by the OL Operator
At the level of gene regulation, phage λ is probably the best-understood organism on Earth. Yet, every time I think we have most of this phage figured out and there is not much else to learn, I read yet another interesting and insightful article about λ.
For those who are not familiar with phage λ genetic circuitry or don’t remember it from genetics class, here is a primer on the basics: The genetic switch controls whether an infecting phage undergoes lytic or lysogenic growth and in phage λ this is centrally controlled by whether or not the cI repressor gene is expressed. The default decision is to undergo lytic growth whereby the phage replicates within the host cell and lyses the cell, releasing progeny, unless repressor is made. CI is expressed from the PRM promoter and there is a divergent, lytic promoter, PR, which drives expression of cro. Cro antagonizes CI and drives lytic growth. These two promoters are both controlled by the operator OR, which is actually three operator sites (OR1, OR2, OR3). CI binds cooperatively to OR1 and OR2, shutting down expression from PR and thus lytic growth. It also activates its own expression at PRM. When concentrations of CI in the cell are sufficient it binds to OR3 and blocks its own transcription, thus maintaining a steady state of CI in the cell. Not only does CI bind to the OR sites cooperatively, but there are similar sites ~2.3 Kbp away at the OL operator that also binds CI. Further cooperativity between CI bound at OR and OL leads to DNA looping, further enhancing the activation and repression of the PRM and PR promoters, respectively. (If this is a bit much, there is a great minireview on λ genetics by Court, Oppenheim and Adhya1 that gives a more in depth explaination)
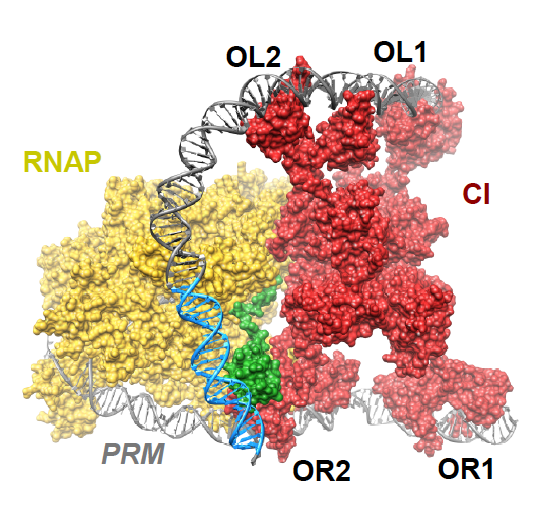
How DNA looping enhances repression of PR is pretty straight forward, but what has not been well understood is how the OL site increases CI activation of PRM. Cui et al. provide support for the “UP contact model” in which OL-OR looping brings an UP element for the PL promoter close enough to PRM so that it can be contacted by the C-terminal domain of the α-subunit of an RNA polymerase bound at PRM. They show that activation requires the UP element, an intact α C-terminal domain and CI-mediated DNA looping, thus confirming the model. Furthermore, they provide structural models supporting the feasibility of contact between the UP element and the α-subunit. The figure above (Fig. 4D in the article) shows an octamer of CI bound at two OR and two OL sites looping the DNA so that the α-subunit (green) of RNA polymerase (yellow) can contact the UP element (blue).
Enhancer elements are quite intriguing genetic elements that can activate transcription at a promoter located several kilobases away. They are widespread, existing in eukaryotes, prokaryotes and phage, yet our understanding of how enhancer-promoter complexes work at a mechanistic level still lacks many details. Cui et al. have uncovered an enhancer-like element in λ that both provides further insight into λ genetic circuitry and a potential model for enhancer elements. What wonders will be revealed about this 48.5 Kbp piece of DNA we call λ in the near future? I can’t wait to find out!
1. Court, D., et al. 2007. A new look at bacteriophage λ genetic networks. 189(2): 298-304.
